
Historical climatic variations and rising atmospheric CO2 concentrations which favor net primary production (NPP) over heterotrophic respiration (RH) may have led to increased ecosystem carbon storage and might account for at least part of the "missing" sink required to balance the historical global carbon budget. To test this hypothesis, we employed a transient global terrestrial carbon cycle model of 0.5° spatial resolution. The model was driven with historical time series of monthly climate and the historical record of changes in atmospheric CO2 concentration. The model's cumulative carbon sink for the period 19001988 accounts for about 68% of the estimated total "missing" sink. For the period 1950
Terrestrial ecosystems exchange a large amount of CO2 with the atmosphere each year. Approximately 15% of the atmospheric C, or 120 Gt C, is assimilated in terrestrial-plant photosynthesis each year, and about an equal amount is returned to the atmosphere as CO2 in plant respiration and the decomposition of plant litter and soil organic matter. The many processes that determine the magnitude of the carbon exchanges between the atmosphere and terrestrial ecosystems are sensitive to climate factors and changes in atmospheric CO2. Time series of the
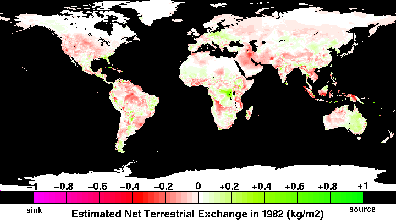
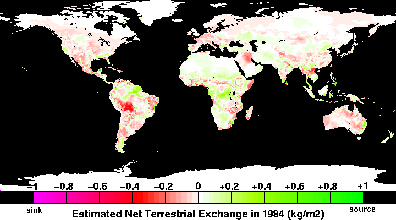
atmospheric CO2
concentration since 1958 show both a long-term secular increase,
largely due to fossil fuel burning, and shorter-term seasonal
oscillations related to the activity of terrestrial biota in the
northern hemisphere. In addition to these regular features, there are
relatively large anomalies that have been attributed to transient
imbalances in natural carbon exchange rates between the atmosphere and
terrestrial ecosystems, perhaps caused by climate fluctuations such as
the El Niño-Southern Oscillation (ENSO) phenomenon. It has been
suggested that subtle and systematic changes in the net carbon fluxes
as the result of climate variations or rising CO2
concentrations over the past century may account for a substantial
portion of the "missing" carbon sink of approximately 1 to 2 Gt C yr-1 needed to balance the contemporary atmospheric CO2 budget.
The rate at which carbon accumulates or is released from terrestrial ecosystems depends on not only the rate of physiological processes but also the size of various plant and soil pools, which have a wide range of turnover times. Changes in carbon storage in relatively slow turnover pools, especially wood and soil humus, determine whether terrestrial systems are net sources or sinks of carbon with respect to the atmosphere (Post et al. 1996). It is therefore necessary to keep track of not only the rates of carbon fixation, respiration and decomposition, and their fluctuation with environmental conditions, but also the size of the living and dead organic matter pools that are affected. Accordingly, we employ a global georeferenced compartment model of carbon in vegetation and soil (King et al. 1997; Post et al. 1997).
We partition the Earth's surface with a square lattice of 0.5 degree latitude by 0.5 degree longitude resolution. Each land cell is assigned to one of 31 ecosystem types according to the vegetation map of Matthews (1983) and to one of 106 soil types according to the FAO Soil Map of the World. For each vegetated cell carbon uptake as net primary production (NPP) is modeled with the Miami Model (Lieth 1975) modified to include a CO2 response term based on theory and observation of leaf photosynthetic response that allows NPP to increase with increasing atmospheric CO2. The calculated NPP is partitioned among four plant parts: leaf, branch, stem, and root. Litter production for each plant part is a function of life span parameters and the biomass of each plant part. The dynamics of litter and soil carbon are calculated with an implementation of the Rothamsted soil carbon turnover model (Jenkinson 1990).
We equilibrated and initialized the compartment model for each vegetated land cell using climate and atmospheric CO2 concentrations appropriate for the end of the nineteenth century. We then simulated changes in NPP and carbon storage from 1900 to 1988 using monthly temperature and precipitation from observed weather records. The history of atmospheric CO2 concentrations was prescribed using CO2 measurements from the Siple Station ice core for years before 1959 and the atmospheric monitoring station at Mauna Loa Observatory thereafter.
Figure 2 shows modeled changes in terrestrial biospheric carbon of natural ecosystems, not significantly affected by land-use. There was a nearly linear increase in vegetation carbon
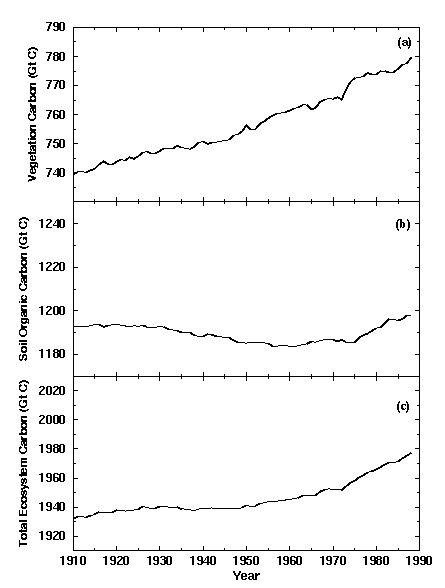
of about 40 Gt C (5%). Since the model's vegetation carbon pools are
not directly affected by either temperature or precipitation, changes
in vegetation carbon are the consequence of changes in NPP (Fig. 3a).
Interannual variability in temperature and precipitation causes
fluctuations in NPP on the order of 1.0 Gt C with occasional changes of
2.0 Gt C and even 3.0 Gt C (e.g., 19721975).
There is evidence of a decline in total soil organic carbon over the period 19101975.
Soil carbon increases after 1975, resulting in a small net gain of 13.6
Gt C (1%) over the entire simulation. The loss of soil carbon is
coincident with declining litter inputs (Fig. 3b), suggesting that the
loss in soil C is a consequence of changes in litter inputs. The loss
in soil carbon slows and then reverses itself as litter production
increases. The rather abrupt increase in modeled soil organic carbon
after 1975 (Fig. 2c) reflects accelerated CO2 enhancement of NPP (Fig. 3a) and an abrupt increase in litter inputs to the soil system (Fig. 3b).
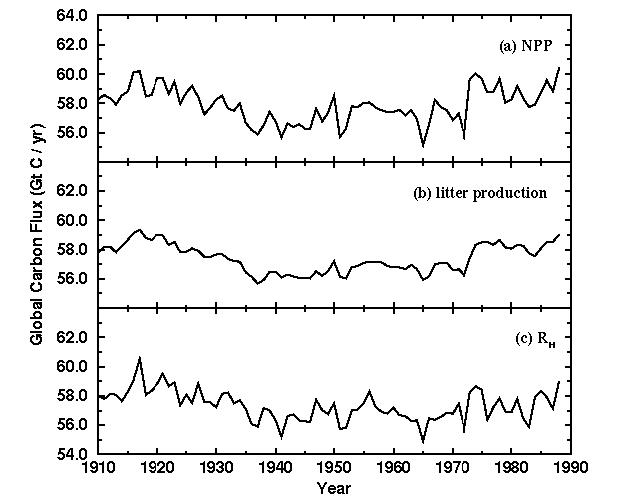
Climate-induced
changes in decomposition rates may contribute to the decline in soil
carbon, but increased decomposition rates during the period of soil
carbon decline are not evident in changes in RH, the carbon released as CO2 during decomposition (Fig. 3c).
Changes in global total ecosystem carbon constitute net terrestrial biospheric exchange with the atmosphere. Our modeled estimate of this net biospheric flux, the difference between RH and NPP, is shown in Fig. 4. Positive values indicate a biospheric source; negative values indicate a biospheric sink. The modeled net biospheric flux shows considerable interannual variability. Interannual differences in the modeled flux are commonly 0.5-1.0 Gt C, with occasional excursions of 2.0 Gt C (Fig. 1). The modeled biosphere frequently changes from source to sink or sink to source over periods of 1 to 3 years.
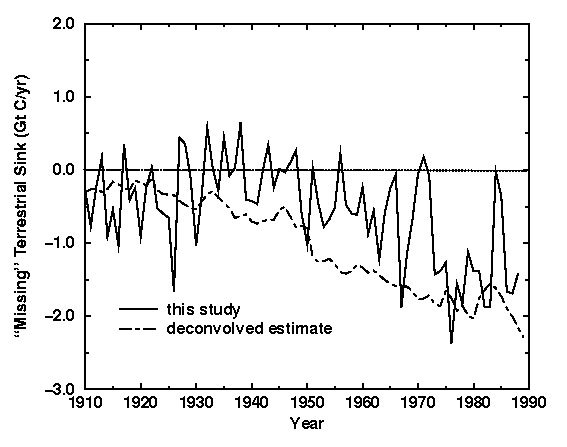
Figure 4 also shows an estimate of the net terrestrial biospheric flux derived by deconvolution of the historical atmospheric CO2 record. The deconvolution generates the residual flux required to balance the observed atmospheric CO2 record with industrial CO2 emissions, net land-use emissions (Houghton and Hackler 1995), and modeled oceanic uptake estimated using a one-dimensional box-diffusion ocean model. Assuming that the residual flux is the result of terrestrial processes, the deconvolution provides an independent estimate of the historical net carbon exchange between the atmosphere and the terrestrial biosphere. In performing a deconvolution, some inputs are necessarily smoothed to prevent large fluctuations that arise from a lack of smoothness resulting from measurement uncertainties. There is no way to unsmooth the deconvolved estimate or smooth the results of the forward calculation of our model that would make them consistent. While this difference complicates our analysis the comparison between model and deconvolution is useful.
There is a general agreement in trends or temporal evolution of the
natural biospheric flux (without land-use) over the 80-year period
compared to the deconvolution. Allowing for interannual variability,
the modeled flux with a land-use flux added agrees reasonably well with
the deconvolution in the period of 1910 to 1930 and in the sink period
of 19751980.
In other periods modeled flux disagrees with the deconvolved estimate.
The model predicts a relatively weak sink and perhaps a source during
the period 1930
1950
when the deconvolution suggests an increasingly stronger sink.
Moreover, the model predicts a sink that is too weak from 1950 to 1975
compared to the deconvolution.
Our simulations provide model evidence that the observed temporal pattern or evolution of net biospheric flux is partially attributable to a combination of climate and atmospheric CO2 forcings and perhaps to climate variation alone. However, the model is unable to reproduce the temporal history and magnitude of the "missing" terrestrial sink derived by deconvolution. Assuming that the deconvolution is an accurate method of determining the historical net biosphere flux, then either the land-use term is too large for most of the period since 1930, or the modeled sink in response to changes in climate and atmospheric CO2 is too small.
References
Houghton, R. A. and J. L. Hackler. 1995. Continental scale estimates of the biotic carbon flux from land cover change: 18501980, ORNL/CDIAC-79, Carbon Dioxide Information Analysis Center, Oak Ridge National Laboratory, Oak Ridge, Tennessee.
Jenkinson, D. S. 1990. The turnover of organic carbon and nitrogen in soil. Phil. Trans. R. Soc. Lond. B 329:361368.
King, A. W., W. M. Post, and S. D. Wullschleger. 1997. The potential
response of terrestrial carbon storage to changes in climate and
atmospheric CO2. Climatic Change 35:199227.
Lieth, H. 1975. Modelling the primary productivity of the world. pp. 237263. In H. Lieth and R. H. Whittaker (eds.), Primary Productivity of the Biosphere. Springer-Verlag, Berlin.
Matthews, E. 1983. Global vegetation and land use: New high resolution
data bases for climate studies. J. Clim. Appl. Met. 22:474487.
Post, W. M., King, A. W., and S. D. Wullschleger 1996. Soil organic
matter models and global estimates of soil organic carbon. pp. 201222.
In P. Smith, J. Smith and D. Powlson (eds.), Evaluation of Soil Organic
Matter Models Using Existing Long-Term Datasets. Springer-Verlag,
Berlin.
Post, W. M., King, A. W., and S. D. Wullschleger. 1997. Historical
variations in terrestrial carbon storage. Global Biogeochemical Cycles
11:99109.
Investigators
Wilfred M. Post
Oak Ridge National Laboratory
P.O. Box 2008
Bldg. 1000, MS-6335
Oak Ridge, Tennessee 37831-6335
(865) 576-3431
FAX: (865) 574-2232
E-mail: wmp@ornl.gov
Anthony W. King
Oak Ridge National Laboratory
P.O. Box 2008
Bldg. 1000, MS-6335
Oak Ridge, Tennessee 37831-6335
(865) 574-3436
FAX: (865) 574-2232
E-mail: awk@ornl.gov
Forrest Hoffman
Oak Ridge National Laboratory
P.O. Box 2008
Bldg. 1505, MS-6036
Oak Ridge, Tennessee 37831-6335
(865) 576-7680
E-mail: forrest@ornl.gov
Stan D. Wullschleger
Oak Ridge National Laboratory
P.O. Box 2008
Bldg. 1059, MS-6335
Oak Ridge, Tennessee 37831-6422
(865) 574-7389
E-mail: wullschlegsd@ornl.gov
Program Manager
Roger C. Dahlman
Office of Health and Environmental Research ER-74,
U.S. Department of Energy
19901 Germantown Road
Germantown, Maryland, D.C. 20874
(301) 903-4951
FAX (301) 903-8519
Email: roger.dahlman@oer.doe.gov